ความเสถียรของแบบจำลองทางคณิตศาสตร์ประเภทนักล่า-เหยื่อ รายวิชา: การศึกษาเชิงคุณภาพของแบบจำลองนักล่า-เหยื่อ
แบบจำลองของโคลโมโกรอฟตั้งสมมติฐานที่สำคัญประการหนึ่ง เนื่องจากสันนิษฐานว่านี่หมายความว่ามีกลไกในประชากรเหยื่อที่ควบคุมจำนวนของมันแม้ว่าจะไม่มีผู้ล่าก็ตาม
น่าเสียดายที่การกำหนดแบบจำลองดังกล่าวไม่อนุญาตให้ตอบคำถามนั้น เมื่อเร็วๆ นี้มีการถกเถียงกันมากมาย ซึ่งเราได้กล่าวไว้แล้วในตอนต้นของบท: ประชากรผู้ล่าจะสามารถมีอิทธิพลด้านกฎระเบียบต่อประชากรเหยื่อเพื่อให้ระบบทั้งหมดยั่งยืนได้อย่างไร ดังนั้น เราจะกลับไปสู่แบบจำลอง (2.1) ซึ่งกลไกการควบคุมตนเอง (เช่น การควบคุมผ่านการแข่งขันภายในความจำเพาะ) หายไปในประชากรที่เป็นเหยื่อ (เช่นเดียวกับในประชากรนักล่า) ดังนั้นกลไกเดียวในการควบคุมจำนวนสายพันธุ์ที่รวมอยู่ในชุมชนคือความสัมพันธ์ทางโภชนาการระหว่างผู้ล่าและเหยื่อ
ในที่นี้ (ดังนั้น ไม่เหมือนกับรุ่นก่อนหน้า ตามธรรมชาติแล้ว วิธีแก้ปัญหา (2.1) ขึ้นอยู่กับประเภทเฉพาะของฟังก์ชันทางโภชนาการ ซึ่งในทางกลับกัน จะถูกกำหนดโดยธรรมชาติของการปล้นสะดม กล่าวคือ กลยุทธ์ทางโภชนาการของนักล่าและกลยุทธ์การป้องกันของ เหยื่อ ฟังก์ชั่นทั่วไปทั้งหมดเหล่านี้ (ดูรูปที่ I) เป็นคุณสมบัติดังต่อไปนี้:
ระบบ (2.1) มีจุดคงที่ที่ไม่น่าสนใจจุดหนึ่ง ซึ่งพิกัดถูกกำหนดจากสมการ
![]()
ภายใต้ข้อจำกัดทางธรรมชาติ
มีจุดคงที่อีกจุดหนึ่ง (0, 0) ซึ่งสอดคล้องกับสมดุลเล็กน้อย มันง่ายที่จะแสดงให้เห็นว่าจุดนี้เป็นจุดอาน และจุดแยกคือแกนพิกัด
สมการคุณลักษณะของจุดมีรูปแบบ
![]()
แน่นอนว่าสำหรับรุ่น Volterra สุดคลาสสิก
ดังนั้นค่าของ f จึงถือได้ว่าเป็นการวัดความเบี่ยงเบนของแบบจำลองที่พิจารณาจากแบบจำลอง Volterra
![]()
จุดที่นิ่งคือโฟกัส และการสั่นปรากฏในระบบ เมื่อความไม่เท่าเทียมกันที่ตรงกันข้ามเป็นที่พอใจ ก็จะมีโหนดและไม่มีการแกว่งในระบบ ความเสถียรของสถานะสมดุลนี้ถูกกำหนดโดยเงื่อนไข
กล่าวคือ มันขึ้นอยู่กับประเภทของหน้าที่ทางโภชนาการของนักล่าอย่างมีนัยสำคัญ
เงื่อนไข (5.5) สามารถตีความได้ดังต่อไปนี้: เพื่อความมั่นคงของความสมดุลที่ไม่เป็นอันตรายของระบบนักล่า-เหยื่อ (และเพื่อการดำรงอยู่ของระบบนี้) ก็เพียงพอแล้วที่ในบริเวณใกล้เคียงของสถานะนี้ สัดส่วนสัมพัทธ์ของเหยื่อที่ใช้ไป โดยผู้ล่าจะเพิ่มขึ้นตามจำนวนเหยื่อที่เพิ่มขึ้น แท้จริงแล้วสัดส่วนของเหยื่อ (จากจำนวนทั้งหมด) ที่นักล่าบริโภคนั้นอธิบายได้ด้วยฟังก์ชันที่หาอนุพันธ์ได้ สภาพที่จะเพิ่มขึ้น (อนุพันธ์เชิงบวก) ดูเหมือนว่า
![]()
เงื่อนไขสุดท้ายที่ทำ ณ จุดนั้นไม่มีอะไรมากไปกว่าเงื่อนไข (5.5) เพื่อความมั่นคงของสมดุล ด้วยความต่อเนื่องก็ต้องเติมเต็มให้ตรงจุดด้วย ดังนั้น หากจำนวนผู้ประสบภัยในละแวกนี้แล้ว
ให้ฟังก์ชันทางโภชนาการ V มีรูปแบบดังแสดงในรูปที่ 1 11, a (คุณลักษณะของสัตว์ไม่มีกระดูกสันหลัง) แสดงว่าสำหรับค่าจำกัดทั้งหมด (เนื่องจากนูนขึ้น)
นั่นคือ สำหรับมูลค่าใดๆ ของจำนวนเหยื่อที่อยู่นิ่ง ความไม่เท่าเทียมกัน (5.5) ไม่เป็นที่น่าพอใจ
ซึ่งหมายความว่าในระบบที่มีฟังก์ชันทางโภชนาการประเภทนี้ จะไม่มีความสมดุลที่ไม่สำคัญที่เสถียร ผลลัพธ์ที่เป็นไปได้หลายประการ: จำนวนเหยื่อและผู้ล่าเพิ่มขึ้นอย่างไม่มีกำหนด หรือ (เมื่อวิถีโคจรผ่านใกล้แกนพิกัดแกนใดแกนหนึ่ง) เนื่องจากเหตุผลแบบสุ่ม จำนวนเหยื่อหรือจำนวนผู้ล่าจะเท่ากับศูนย์ หากเหยื่อตาย ผู้ล่าก็จะตายหลังจากนั้นระยะหนึ่ง แต่ถ้าผู้ล่าตายก่อน จำนวนเหยื่อก็จะเริ่มเพิ่มขึ้นแบบทวีคูณ ตัวเลือกที่สาม - การเกิดขึ้นของวงจรขีดจำกัดที่มั่นคง - เป็นไปไม่ได้ ซึ่งได้รับการพิสูจน์อย่างง่ายดาย
ที่จริงแล้วการแสดงออก

ในจตุภาคบวกจะเป็นค่าบวกเสมอ เว้นแต่จะมีรูปแบบดังแสดงในรูปที่ 1 11 ก. จากนั้น ตามเกณฑ์ Dulac ไม่มีวิถีแบบปิดในภูมิภาคนี้ และไม่สามารถมีวงจรจำกัดที่มั่นคงได้
ดังนั้นเราสามารถสรุปได้ว่า: ถ้าฟังก์ชันทางโภชนาการมีรูปแบบดังแสดงในรูปที่ 1 11 จากนั้นผู้ล่าไม่สามารถเป็นตัวควบคุมที่รับประกันความเสถียรของประชากรเหยื่อและด้วยเหตุนี้จึงทำให้มีความเสถียรของทั้งระบบโดยรวม ระบบจะมีเสถียรภาพได้ก็ต่อเมื่อประชากรที่เป็นเหยื่อมีกลไกการกำกับดูแลภายในของตัวเอง เช่น การแข่งขันภายในความจำเพาะหรือสัตว์ในสัตว์หลายชนิด ตัวเลือกกฎระเบียบนี้ได้มีการกล่าวถึงแล้วใน§§ 3, 4
ก่อนหน้านี้มีการตั้งข้อสังเกตว่าหน้าที่ทางโภชนาการประเภทนี้เป็นลักษณะของผู้ล่าแมลงซึ่ง "เหยื่อ" มักเป็นแมลงเช่นกัน ในทางกลับกัน การสังเกตพลวัตของชุมชนธรรมชาติหลายประเภทประเภท "นักล่า-เหยื่อ" รวมถึงแมลงสายพันธุ์ต่างๆ แสดงให้เห็นว่าพวกมันมีลักษณะเฉพาะจากการผันผวนของแอมพลิจูดที่มีขนาดใหญ่มากและเป็นประเภทที่เฉพาะเจาะจงมาก
โดยปกติแล้วหลังจากที่ตัวเลขเพิ่มขึ้นทีละน้อยไม่มากก็น้อย (ซึ่งอาจเกิดขึ้นได้ทั้งแบบซ้ำซากหรือในรูปแบบของการแกว่งที่มีแอมพลิจูดเพิ่มขึ้น) การลดลงอย่างรวดเร็วจะเกิดขึ้น (รูปที่ 14) จากนั้นภาพจะเกิดซ้ำ เห็นได้ชัดว่าธรรมชาติของพลวัตของจำนวนแมลงชนิดนี้สามารถอธิบายได้ด้วยความไม่แน่นอนของระบบนี้ที่จำนวนต่ำและปานกลาง และการกระทำของหน่วยงานกำกับดูแลที่มีประสิทธิภาพในจำนวนแมลงจำนวนมาก

ข้าว. 14. พลวัตของประชากรของ Cardiaspina albitextura Psyllid ของออสเตรเลียที่กินบนต้นยูคาลิปตัส (จากบทความ: Clark L. R. The Population Dynamics of Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, No. 3, p. 362-380.)
หากระบบ "นักล่า-เหยื่อ" รวมสายพันธุ์ที่มีพฤติกรรมค่อนข้างซับซ้อน (เช่น ผู้ล่าสามารถเรียนรู้ได้หรือเหยื่อสามารถหาที่หลบภัยได้) ความสมดุลที่ไม่สำคัญและมั่นคงก็อาจมีอยู่ในระบบดังกล่าว ข้อความนี้ได้รับการพิสูจน์แล้วค่อนข้างง่าย
ในความเป็นจริง ฟังก์ชั่นทางโภชนาการควรมีรูปแบบดังแสดงในรูปที่ 1 11 ค. จุดบนกราฟนี้คือจุดสัมผัสของเส้นตรงที่ดึงมาจากจุดกำเนิดของกราฟฟังก์ชันโภชนาการ แน่นอนว่า ณ จุดนี้ฟังก์ชันจะมีค่าสูงสุด นอกจากนี้ยังเป็นเรื่องง่ายที่จะแสดงให้เห็นว่าเงื่อนไข (5.5) เป็นที่พอใจสำหรับทุกคน ดังนั้น ความสมดุลที่ไม่เป็นอันตรายซึ่งจำนวนเหยื่อน้อยลงจะคงที่แบบไม่แสดงอาการ
อย่างไรก็ตาม เราไม่สามารถพูดอะไรได้ว่าขอบเขตเสถียรภาพของสมดุลนี้มีขนาดใหญ่เพียงใด ตัวอย่างเช่น หากมีรอบขีดจำกัดที่ไม่เสถียร ขอบเขตนี้จะต้องอยู่ภายในวงจรนั้น หรือทางเลือกอื่น: สมดุลที่ไม่น่าสนใจ (5.2) ไม่เสถียร แต่มีวงจรขีดจำกัดที่มั่นคง ในกรณีนี้เราสามารถพูดคุยเกี่ยวกับความเสถียรของระบบนักล่าและเหยื่อได้ ตั้งแต่นิพจน์ (5.7) เมื่อเลือกฟังก์ชันทางโภชนาการเช่นรูปที่ 11 ใน สามารถเปลี่ยนเครื่องหมายได้เมื่อเปลี่ยนที่ ดังนั้นเกณฑ์ Dulac จะใช้ไม่ได้ที่นี่ และคำถามเกี่ยวกับการมีอยู่ของวงจรขีดจำกัดยังคงเปิดอยู่

ตามข้อตกลงลงวันที่ ___.___, 20___ เกี่ยวกับการให้บริการการศึกษาแบบชำระเงิน
กระทรวงศึกษาธิการและวิทยาศาสตร์ สหพันธรัฐรัสเซีย
สาขาลีสเวนสกี้
รัฐระดับการใช้งาน มหาวิทยาลัยเทคนิค
ภาควิชาเศรษฐศาสตร์
ในสาขาวิชา "การสร้างแบบจำลองระบบ"
หัวข้อ: ระบบล่าเหยื่อ
สมบูรณ์:
นักเรียน ก. บีไอวีที-06
------------------
ตรวจสอบโดยอาจารย์:
เชสตาคอฟ เอ.พี.
ลิสวา, 2010
เรียงความ
การปล้นสะดมเป็นความสัมพันธ์ทางโภชนาการระหว่างสิ่งมีชีวิตโดยหนึ่งในนั้น (ผู้ล่า) โจมตีอีกตัว (เหยื่อ) และกินส่วนต่าง ๆ ของร่างกายซึ่งก็คือมักจะมีการฆ่าเหยื่อ การปล้นสะดมนั้นตรงกันข้ามกับการกินซากศพ (การตัดศพ) และผลิตภัณฑ์อินทรีย์ที่ย่อยสลาย (detritophagy)
คำจำกัดความอีกประการหนึ่งของการปล้นสะดมก็เป็นที่นิยมเช่นกัน ซึ่งเสนอว่ามีเพียงสิ่งมีชีวิตที่กินสัตว์เท่านั้นที่จะเรียกว่าผู้ล่า ตรงกันข้ามกับสัตว์กินพืชที่กินพืช
นอกจากสัตว์หลายเซลล์แล้ว กลุ่มผู้ประท้วง เชื้อรา และพืชชั้นสูงก็สามารถทำหน้าที่เป็นผู้ล่าได้
ขนาดประชากรของผู้ล่าส่งผลกระทบต่อขนาดประชากรของเหยื่อและในทางกลับกัน แบบจำลองทางคณิตศาสตร์ Lotka-Volterra อธิบายพลวัตของประชากร อย่างไรก็ตาม โมเดลนี้เป็นนามธรรมในระดับสูง และไม่ได้อธิบายความสัมพันธ์ที่แท้จริงระหว่างผู้ล่าและเหยื่อ และถือได้ว่าเป็นระดับแรกของการประมาณค่านามธรรมทางคณิตศาสตร์เท่านั้น
ในกระบวนการวิวัฒนาการร่วมกัน ผู้ล่าและเหยื่อจะปรับตัวเข้าหากัน ผู้ล่าปรากฏตัวและพัฒนาวิธีการตรวจจับและโจมตี ส่วนเหยื่อก็มีวิธีการรักษาความลับและการป้องกัน นั่นเป็นเหตุผล ความเสียหายที่ยิ่งใหญ่ที่สุดเหยื่ออาจได้รับอันตรายจากผู้ล่ารายใหม่ที่พวกเขายังไม่ได้เข้าร่วม "การแข่งขันทางอาวุธ"
ผู้ล่าสามารถเชี่ยวชาญในการใช้เหยื่อหนึ่งสายพันธุ์หรือมากกว่านั้น ซึ่งทำให้พวกมันประสบความสำเร็จในการล่าสัตว์โดยเฉลี่ยมากขึ้น แต่ต้องพึ่งพาสายพันธุ์เหล่านั้นมากขึ้น
ระบบล่าเหยื่อ
ปฏิสัมพันธ์ระหว่างผู้ล่าและเหยื่อเป็นความสัมพันธ์แนวดิ่งประเภทหลักระหว่างสิ่งมีชีวิต ซึ่งสสารและพลังงานถูกถ่ายโอนผ่านห่วงโซ่อาหาร
สมดุลของ V. x - และ. สำเร็จได้ง่ายที่สุดถ้า ห่วงโซ่อาหารมีลิงก์อย่างน้อยสามลิงก์ (เช่น หญ้า - นา - สุนัขจิ้งจอก) ในเวลาเดียวกัน ความหนาแน่นของประชากรไฟโตฟาจถูกควบคุมโดยความสัมพันธ์กับการเชื่อมโยงทั้งด้านล่างและด้านบนของห่วงโซ่อาหาร
ขึ้นอยู่กับลักษณะของเหยื่อและประเภทของนักล่า (จริง, สัตว์กินหญ้า), การพึ่งพาพลวัตของประชากรที่แตกต่างกันนั้นเป็นไปได้ ยิ่งไปกว่านั้น ภาพยังมีความซับซ้อนเนื่องจากความจริงที่ว่าผู้ล่านั้นไม่ค่อยมีสัตว์ประเภทเดียวมากนัก (เช่น กินเหยื่อประเภทเดียว) บ่อยครั้งเมื่อประชากรของเหยื่อประเภทหนึ่งหมดลงและการจับมันต้องใช้ความพยายามมากเกินไป ผู้ล่าจึงเปลี่ยนไปใช้เหยื่อประเภทอื่น นอกจากนี้ประชากรเหยื่อหนึ่งกลุ่มสามารถถูกเอารัดเอาเปรียบโดยผู้ล่าหลายสายพันธุ์
ด้วยเหตุนี้ ผลของการทำให้ขนาดประชากรของเหยื่อเต้นเป็นจังหวะ ซึ่งมักอธิบายไว้ในวรรณกรรมด้านสิ่งแวดล้อม ตามมาด้วยขนาดประชากรของเหยื่อที่เต้นเป็นจังหวะด้วยความล่าช้าบางอย่าง จึงเป็นสิ่งที่หาได้ยากในธรรมชาติ
ความสมดุลระหว่างผู้ล่าและเหยื่อในสัตว์นั้นได้รับการดูแลโดยกลไกพิเศษที่ป้องกันการกำจัดเหยื่อโดยสมบูรณ์ ดังนั้นเหยื่อสามารถ:
- วิ่งหนีจากนักล่า (ในกรณีนี้อันเป็นผลมาจากการแข่งขันความคล่องตัวของทั้งเหยื่อและผู้ล่าเพิ่มขึ้นซึ่งเป็นเรื่องปกติโดยเฉพาะอย่างยิ่งสำหรับสัตว์บริภาษที่ไม่มีที่ซ่อนจากผู้ไล่ตาม)
- ได้รับ ภาพวาดป้องกัน (<притворяться>ใบไม้หรือกิ่งไม้) หรือในทางกลับกันสีสดใส (เช่นสีแดง) เตือนนักล่าเกี่ยวกับรสขม
- ซ่อนตัวอยู่ในที่พักอาศัย
- ย้ายไปใช้มาตรการป้องกันเชิงรุก (สัตว์กินพืชมีเขา ปลามีหนาม) มักจะข้อต่อ (นกล่าเหยื่อรวมตัวกันขับไล่ว่าว กวางตัวผู้ และไซกัสเข้ายึดครอง<круговую оборону>จากหมาป่า ฯลฯ)
บ่อยครั้งสมาชิกของสปีชีส์หนึ่ง (ประชากร) กินสมาชิกของสปีชีส์อื่นเป็นอาหาร
แบบจำลอง Lotka-Volterra เป็นแบบจำลองของการดำรงอยู่ร่วมกันของประชากรสองคนประเภท "นักล่า - เหยื่อ"
แบบจำลองนักล่า-เหยื่อได้รับการพัฒนาครั้งแรกโดย A. Lotka ในปี 1925 ซึ่งใช้แบบจำลองนี้เพื่ออธิบายพลวัตของการมีปฏิสัมพันธ์ระหว่างประชากรทางชีววิทยา ในปี 1926 โดยเป็นอิสระจาก Lotka แบบจำลองที่คล้ายกัน (และซับซ้อนกว่า) ได้รับการพัฒนาโดยนักคณิตศาสตร์ชาวอิตาลี V. Volterra ซึ่งมีการวิจัยเชิงลึกในสาขานี้ ปัญหาสิ่งแวดล้อมวางรากฐาน ทฤษฎีทางคณิตศาสตร์ชุมชนทางชีววิทยาหรือที่เรียกว่า นิเวศวิทยาทางคณิตศาสตร์
ใน รูปแบบทางคณิตศาสตร์ระบบสมการที่เสนอมีรูปแบบดังนี้
โดยที่ x คือจำนวนเหยื่อ y คือจำนวนผู้ล่า t คือเวลา α, β, γ, δ คือสัมประสิทธิ์ที่สะท้อนถึงปฏิสัมพันธ์ระหว่างประชากร
การกำหนดปัญหา
พิจารณาพื้นที่ปิดซึ่งมีประชากรสองกลุ่ม ได้แก่ สัตว์กินพืช (“เหยื่อ”) และผู้ล่า เชื่อกันว่าไม่มีการนำเข้าหรือส่งออกสัตว์และมีอาหารเพียงพอสำหรับสัตว์กินพืช จากนั้นสมการการเปลี่ยนแปลงจำนวนเหยื่อ (เฉพาะเหยื่อ) จะอยู่ในรูปแบบ:
โดยที่ $α$ คืออัตราการเกิดของเหยื่อ
$x$ คือขนาดของประชากรเหยื่อ
$\frac(dx)(dt)$ คืออัตราการเติบโตของประชากรเหยื่อ
เมื่อผู้ล่าไม่ล่า พวกมันอาจสูญพันธุ์ได้ ซึ่งหมายความว่าสมการสำหรับจำนวนผู้ล่า (เฉพาะผู้ล่า) จะกลายเป็น:
โดยที่ $γ$ คืออัตราการสูญเสียของผู้ล่า
$y$ คือขนาดของประชากรนักล่า
$\frac(dy)(dt)$ คืออัตราการเติบโตของประชากรนักล่า
เมื่อผู้ล่าและเหยื่อพบกัน (ความถี่ของการประชุมเป็นสัดส่วนโดยตรงกับผลิตภัณฑ์) ผู้ล่าจะทำลายเหยื่อด้วยค่าสัมประสิทธิ์ ผู้ล่าที่ได้รับอาหารอย่างดีสามารถสืบพันธุ์ลูกหลานโดยมีค่าสัมประสิทธิ์ ดังนั้นระบบสมการของแบบจำลองจะอยู่ในรูปแบบ:
การแก้ปัญหา
เรามาสร้างแบบจำลองทางคณิตศาสตร์ของการอยู่ร่วมกันของประชากรทางชีววิทยาสองคนประเภท "นักล่า-เหยื่อ" กัน
ปล่อยให้ประชากรทางชีววิทยาสองคนอาศัยอยู่ร่วมกันในสภาพแวดล้อมที่แยกจากกัน สภาพแวดล้อมคงที่และจัดให้มี ปริมาณไม่จำกัดทุกสิ่งที่จำเป็นสำหรับชีวิตเป็นประเภทหนึ่ง - การเสียสละ อีกสายพันธุ์หนึ่ง - ผู้ล่า - อาศัยอยู่ในสภาวะนิ่ง แต่กินเหยื่อเท่านั้น แมว หมาป่า หอก สุนัขจิ้งจอกสามารถทำหน้าที่เป็นผู้ล่าได้ ส่วนไก่ กระต่าย ปลาคาร์พไม้กางเขน และหนูสามารถทำหน้าที่เป็นเหยื่อได้ ตามลำดับ
หากจะให้เจาะจง ให้ถือว่าแมวเป็นผู้ล่า และไก่เป็นเหยื่อ
ดังนั้น ไก่และแมวจึงอาศัยอยู่ในพื้นที่ห่างไกล เช่น ลานฟาร์ม สภาพแวดล้อมให้อาหารไก่ไม่จำกัด ส่วนแมวกินเฉพาะไก่เท่านั้น ให้เราแสดงโดย
$x$ – จำนวนไก่
$у$ – จำนวนแมว
เมื่อเวลาผ่านไป จำนวนไก่และแมวเปลี่ยนแปลงไป แต่เราจะถือว่า $x$ และ $y$ เป็นฟังก์ชันต่อเนื่องของเวลา t ลองเรียกตัวเลขคู่หนึ่งว่า $x, y)$ สถานะของโมเดล
มาดูกันว่าสถานะของโมเดล $(x, y).$ เปลี่ยนแปลงไปอย่างไร
ลองพิจารณา $\frac(dx)(dt)$ – อัตราการเปลี่ยนแปลงของจำนวนไก่
หากไม่มีแมว จำนวนไก่ก็จะเพิ่มมากขึ้น และยิ่งเร็วเท่าไรก็ยิ่งมีจำนวนไก่มากขึ้นเท่านั้น เราจะถือว่าการพึ่งพานั้นเป็นเส้นตรง:
$\frac(dx)(dt) a_1 x$,
$a_1$ เป็นค่าสัมประสิทธิ์ที่ขึ้นอยู่กับสภาพความเป็นอยู่ของไก่ อัตราตายตามธรรมชาติ และอัตราการเกิดของไก่เท่านั้น
$\frac(dy)(dt)$ – อัตราการเปลี่ยนแปลงของจำนวนแมว (หากไม่มีไก่) ขึ้นอยู่กับจำนวนแมว y
หากไม่มีไก่ จำนวนแมวก็จะลดลง (ไม่มีอาหาร) และพวกมันก็จะตายไป เราจะถือว่าการพึ่งพานั้นเป็นเส้นตรง:
$\frac(dy)(dt) - a_2 y$.
ในระบบนิเวศ อัตราการเปลี่ยนแปลงจำนวนของแต่ละสายพันธุ์จะถือเป็นสัดส่วนกับปริมาณของมันด้วย แต่จะมีค่าสัมประสิทธิ์เท่านั้นที่ขึ้นอยู่กับจำนวนบุคคลในสายพันธุ์อื่น ดังนั้น สำหรับไก่ ค่าสัมประสิทธิ์นี้จะลดลงตามจำนวนแมวที่เพิ่มขึ้น และสำหรับแมว ค่าสัมประสิทธิ์นี้จะเพิ่มขึ้นตามจำนวนไก่ที่เพิ่มขึ้น เราจะถือว่าการพึ่งพานั้นเป็นเส้นตรงด้วย จากนั้นเราจะได้ระบบ สมการเชิงอนุพันธ์:
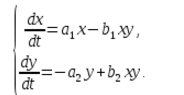
ระบบสมการนี้เรียกว่าแบบจำลองโวลแตร์รา-ลอตกา
a1, a2, b1, b2 – ค่าสัมประสิทธิ์ตัวเลข ซึ่งเรียกว่าพารามิเตอร์โมเดล
อย่างที่คุณเห็นลักษณะของการเปลี่ยนแปลงสถานะของโมเดล (x, y) จะถูกกำหนดโดยค่าของพารามิเตอร์ ด้วยการเปลี่ยนพารามิเตอร์เหล่านี้และการแก้ระบบสมการแบบจำลอง ทำให้สามารถศึกษารูปแบบของการเปลี่ยนแปลงในสถานะของระบบนิเวศได้
การใช้โปรแกรม MATLAB ระบบสมการ Lotka-Volterra ได้รับการแก้ไขดังนี้
ในรูป ภาพที่ 1 แสดงวิธีแก้ปัญหาของระบบ วิธีแก้ปัญหาจะแตกต่างกันไปขึ้นอยู่กับเงื่อนไขเริ่มต้น สีที่ต่างกันวิถี
ในรูป 2 นำเสนอวิธีแก้ปัญหาเดียวกัน แต่คำนึงถึงแกนเวลา t (นั่นคือ มีการขึ้นอยู่กับเวลา)
ย้อนกลับไปในยุค 20 A. Lotka และต่อมา V. Volterra เป็นอิสระจากเขา เสนอแบบจำลองทางคณิตศาสตร์ที่อธิบายความผันผวนของคอนจูเกตในประชากรของนักล่าและเหยื่อ ลองพิจารณารุ่น Lotka-Volterra รุ่นที่ง่ายที่สุด แบบจำลองนี้ขึ้นอยู่กับสมมติฐานหลายประการ:
1) ประชากรเหยื่อในกรณีที่ไม่มีนักล่าเพิ่มขึ้นแบบทวีคูณ
2) ความกดดันของผู้ล่าขัดขวางการเติบโตนี้
3) การตายของเหยื่อแปรผันตามความถี่ของการเผชิญหน้าระหว่างผู้ล่าและเหยื่อ (หรืออย่างอื่น แปรผันตามผลคูณของความหนาแน่นของประชากร)
4) อัตราการเกิดของนักล่าขึ้นอยู่กับความรุนแรงของการบริโภคเหยื่อ
อัตราการเปลี่ยนแปลงขนาดประชากรเหยื่อทันทีสามารถแสดงได้ด้วยสมการ
dN f /dt = r 1 N f - p 1 N f N x,
ที่ไหน r 1 - อัตราการเติบโตของประชากรของเหยื่อในทันทีโดยเฉพาะ p 1 - การเชื่อมต่อการตายของเหยื่อกับความหนาแน่นของนักล่าอย่างต่อเนื่อง เอ็น และ เอ็น เอ็กซ์ - ความหนาแน่นของเหยื่อและผู้ล่าตามลำดับ
อัตราการเติบโตทันทีของประชากรนักล่าในแบบจำลองนี้ถือว่าเท่ากับความแตกต่างระหว่างอัตราการเกิดและการตายคงที่:
dN x /dt = p 2 N x N x – d 2 N x,
ที่ไหนหน้า 2 - ค่าคงที่ที่เกี่ยวข้องกับอัตราการเกิดในประชากรนักล่ากับความหนาแน่นของเหยื่อ a d 2 - อัตราการตายของผู้ล่าโดยเฉพาะ
ตามสมการข้างต้น แต่ละประชากรที่มีปฏิสัมพันธ์ในการเพิ่มขึ้นจะถูกจำกัดโดยประชากรอื่นๆ เท่านั้น เช่น การเพิ่มขึ้นของจำนวนเหยื่อถูกจำกัดด้วยแรงกดดันของผู้ล่า และการเพิ่มจำนวนของผู้ล่านั้นถูกจำกัดด้วยจำนวนเหยื่อที่ไม่เพียงพอ ไม่มีการจำกัดจำนวนประชากรด้วยตนเอง เชื่อกันว่ามีอาหารเพียงพอสำหรับเหยื่อเสมอ ไม่คาดว่าประชากรเหยื่อจะหนีจากการควบคุมของนักล่า แม้ว่าในความเป็นจริงสิ่งนี้จะเกิดขึ้นค่อนข้างบ่อยก็ตาม
แม้จะมีรูปแบบทั่วไปของแบบจำลอง Lotka-Volterra แต่ก็สมควรได้รับความสนใจหากเพียงเพราะมันแสดงให้เห็นว่าแม้แต่ระบบปฏิสัมพันธ์ในอุดมคติระหว่างประชากรทั้งสองก็สามารถก่อให้เกิดพลวัตที่ค่อนข้างซับซ้อนของตัวเลขได้ การแก้ระบบสมการเหล่านี้ช่วยให้เราสามารถกำหนดเงื่อนไขในการรักษาจำนวนคงที่ (สมดุล) ของแต่ละสปีชีส์ได้ ประชากรเหยื่อจะคงที่หากความหนาแน่นของผู้ล่าเท่ากับ r 1 /p 1 และเพื่อให้ประชากรนักล่าคงที่ ความหนาแน่นของเหยื่อจะต้องเท่ากับ d 2 /p 2 หากเราวาดความหนาแน่นของเหยื่อบนแกน x เอ็น และ , และตามลำดับ - ความหนาแน่นของนักล่า เอ็น เอ็กซ์จากนั้นเส้นไอโซไลน์ที่แสดงสภาวะคงที่ของผู้ล่าและเหยื่อจะเป็นเส้นตรงสองเส้นตั้งฉากกันและกับแกนพิกัด (รูปที่ 6, ก) สันนิษฐานว่าความหนาแน่นของเหยื่อต่ำกว่าค่าที่กำหนด (เท่ากับ d 2 /p 2) ความหนาแน่นของนักล่าจะลดลงเสมอและสูงกว่านั้นจะเพิ่มขึ้นเสมอ ดังนั้นความหนาแน่นของเหยื่อจะเพิ่มขึ้นหากความหนาแน่นของนักล่าต่ำกว่าค่าเท่ากับ r 1 / p 1 และลดลงหากอยู่เหนือค่านี้ จุดตัดของเส้นไอโซไลน์สอดคล้องกับสภาพของความอุดมสมบูรณ์ของผู้ล่าและเหยื่ออย่างต่อเนื่อง และจุดอื่นๆ บนระนาบของกราฟนี้เคลื่อนที่ไปตามวิถีปิด ซึ่งสะท้อนถึงความผันผวนอย่างสม่ำเสมอในความอุดมสมบูรณ์ของผู้ล่าและเหยื่อ (รูปที่ 6, ข)ช่วงของการสั่นถูกกำหนดโดยอัตราส่วนเริ่มต้นของความหนาแน่นของผู้ล่าและเหยื่อ ยิ่งใกล้กับจุดตัดของไอโซไลน์ วงกลมก็จะเล็กลงตามเวกเตอร์ และด้วยเหตุนี้ แอมพลิจูดของการแกว่งก็จะน้อยลงตามไปด้วย
ข้าว. 6. การแสดงออกแบบกราฟิกของแบบจำลอง Lotka-Voltaire สำหรับระบบล่าเหยื่อ
หนึ่งในความพยายามครั้งแรกเพื่อให้ได้ความผันผวนของจำนวนผู้ล่าและเหยื่อในการทดลองในห้องปฏิบัติการเป็นของ G.F. เกาส์. วัตถุของการทดลองเหล่านี้คือพารามีเซียมซิลิเอต (พารามีเซียม หาง) และดิดิเนียมซิลิเอตนักล่า (ดิดิเนียม นสุทัม). อาหารของพารามีเซียมคือการเติมแบคทีเรียแขวนลอยลงในอาหารเป็นประจำ และดิดิเนียมกินเฉพาะพารามีเซียมเท่านั้น ระบบนี้กลับกลายเป็นว่าไม่เสถียรอย่างยิ่ง: ความกดดันของนักล่าเมื่อจำนวนของมันเพิ่มขึ้นนำไปสู่การกำจัดเหยื่อโดยสิ้นเชิงหลังจากนั้นประชากรของนักล่าก็ตายไป ทำให้การทดลองซับซ้อนขึ้น Gause ได้จัดที่พักพิงสำหรับเหยื่อโดยนำใยแก้วเล็กน้อยเข้าไปในหลอดทดลองที่มีซิลิเอต ปารามีเซียสามารถเคลื่อนไหวได้อย่างอิสระท่ามกลางเส้นด้ายสำลี แต่ดิดิเนียมทำไม่ได้ ในการทดลองเวอร์ชันนี้ ดิดิเนียมกินพารามีเซียทั้งหมดที่ลอยอยู่ในส่วนที่ไม่มีสำลีของหลอดทดลองและตายไป จากนั้นประชากรพารามีเซียก็กลับคืนมาเนื่องจากการแพร่พันธุ์ของบุคคลที่รอดชีวิตในสถานสงเคราะห์ เกาส์สามารถบรรลุความผันผวนของจำนวนผู้ล่าและเหยื่อได้ก็ต่อเมื่อเขาแนะนำทั้งเหยื่อและผู้ล่าเข้าสู่วัฒนธรรมเป็นครั้งคราว ซึ่งเป็นการจำลองการอพยพ
40 ปีหลังจากงานของเกาส์ การทดลองของเขาถูกทำซ้ำโดยแอล. ลัคคินบิล ซึ่งใช้ซิลิเอตเป็นเหยื่อ พารามีเซียม ออเรเลีย, และในฐานะนักล่าเช่นเดียวกัน ดิดิเนียม นสุทัม. Luckinbill สามารถจัดการความผันผวนของจำนวนประชากรเหล่านี้ได้หลายรอบ แต่เฉพาะในกรณีที่ความหนาแน่นของพารามีเซียมถูกจำกัดด้วยการขาดอาหาร (แบคทีเรีย) และเมทิลเซลลูโลสถูกเติมลงในของเหลวทางวัฒนธรรม - สารที่ช่วยลด ความเร็วในการเคลื่อนที่ของทั้งผู้ล่าและเหยื่อจึงลดความถี่ในการประชุมที่เป็นไปได้ ปรากฎว่าการสั่นระหว่างผู้ล่าและเหยื่อทำได้ง่ายกว่าหากปริมาตรของภาชนะทดลองเพิ่มขึ้น แม้ว่าในกรณีนี้เงื่อนไขของการจำกัดอาหารของเหยื่อก็จำเป็นเช่นกัน หากมีการเพิ่มอาหารส่วนเกินเข้าไปในระบบของผู้ล่าและเหยื่อที่อยู่ร่วมกันในโหมดการสั่น การตอบสนองก็คือจำนวนเหยื่อที่เพิ่มขึ้นอย่างรวดเร็ว ตามมาด้วยจำนวนผู้ล่าที่เพิ่มขึ้น ซึ่งนำไปสู่การกำจัดโดยสิ้นเชิง ประชากรเหยื่อ
แบบจำลอง Lotka และ Volterra ทำหน้าที่เป็นแรงผลักดันในการพัฒนาแบบจำลองอื่นๆ ของระบบล่าเหยื่อที่สมจริงยิ่งขึ้น โดยเฉพาะอย่างยิ่งแบบจำลองกราฟิกที่ค่อนข้างเรียบง่ายที่วิเคราะห์อัตราส่วนของไอโซไลน์ของเหยื่อที่แตกต่างกัน นักล่า ถูกเสนอโดย M. Rosenzweig และ R. MacArthur (Rosenzweig, MacArthur) ตามที่ผู้เขียนเหล่านี้เครื่องเขียน ( = ค่าคงที่) จำนวนเหยื่อในแกนพิกัดของความหนาแน่นของผู้ล่าและเหยื่อสามารถแสดงในรูปแบบของไอโซไลน์นูน (รูปที่ 7, a) จุดหนึ่งที่เส้นไอโซไลน์ตัดกับเส้นความหนาแน่นของเหยื่อสอดคล้องกับความหนาแน่นของเหยื่อขั้นต่ำที่อนุญาต (ประชากรด้านล่างมีความเสี่ยงสูงมากที่จะสูญพันธุ์ หากเพียงเพราะความถี่ของการพบกันระหว่างตัวผู้และตัวเมียต่ำ) และอีกจุดหนึ่ง สอดคล้องกับค่าสูงสุดที่กำหนดโดยปริมาณอาหารที่มีอยู่หรือลักษณะพฤติกรรมของเหยื่อเอง ให้เราย้ำว่าเรายังคงพูดถึงความหนาแน่นขั้นต่ำและสูงสุดในกรณีที่ไม่มีผู้ล่า เมื่อนักล่าปรากฏตัวและจำนวนของมันเพิ่มขึ้นความหนาแน่นของเหยื่อขั้นต่ำที่อนุญาตควรสูงขึ้นอย่างเห็นได้ชัดและค่าสูงสุด - ต่ำกว่า แต่ละค่าของความหนาแน่นของเหยื่อจะต้องสอดคล้องกับความหนาแน่นของนักล่าที่แน่นอนซึ่งทำให้ประชากรเหยื่อมีความคงที่ ตำแหน่งทางเรขาคณิตของจุดดังกล่าวคือไอโซไลน์ของเหยื่อในพิกัดความหนาแน่นของผู้ล่าและเหยื่อ เวกเตอร์ที่แสดงทิศทางของการเปลี่ยนแปลงความหนาแน่นของเหยื่อ (ในแนวนอน) มีทิศทางที่แตกต่างกันในด้านต่างๆ ของไอโซไคลน์ (รูปที่ 7a)

ข้าว. 7. ไอโซลินของประชากรเหยื่อ (a) และผู้ล่า (b) ที่อยู่นิ่ง
สำหรับนักล่านั้น ไอโซไลน์ก็ถูกสร้างขึ้นในพิกัดเดียวกันซึ่งสอดคล้องกับสถานะคงที่ของประชากร เวกเตอร์ที่แสดงทิศทางการเปลี่ยนแปลงของความอุดมสมบูรณ์ของสัตว์นักล่าจะถูกวางทิศทางขึ้นหรือลง ขึ้นอยู่กับด้านของไอโซไลน์ที่พวกมันอยู่ รูปร่างของไอโซไลน์นักล่าดังแสดงในรูปที่ 1 7, ข.ถูกกำหนดโดยประการแรกโดยการปรากฏตัวของเหยื่อที่มีความหนาแน่นขั้นต่ำซึ่งเพียงพอที่จะรักษาจำนวนประชากรของนักล่า (ที่ความหนาแน่นของเหยื่อที่ต่ำกว่านักล่าจะไม่สามารถเพิ่มจำนวนได้) และประการที่สองโดยการมีอยู่ของ ความหนาแน่นสูงสุดของนักล่านั้นเอง ซึ่งเกินกว่าจำนวนนั้นจะลดลงอย่างเป็นอิสระจากจำนวนเหยื่อที่มีอยู่มากมาย

ข้าว. 8. การเกิดขึ้นของโหมดการสั่นในระบบล่าเหยื่อขึ้นอยู่กับตำแหน่งของไอโซไลน์ของผู้ล่าและเหยื่อ
เมื่อรวมเหยื่อและผู้ล่าเข้าด้วยกันบนกราฟเดียว จะมีตัวเลือกที่แตกต่างกันสามแบบ (รูปที่ 8) ถ้าไอโซไลน์ของนักล่าตัดกับไอโซไลน์ของเหยื่อในตำแหน่งที่มันลดลงแล้ว (ที่เหยื่อมีความหนาแน่นสูง) พาหะที่แสดงการเปลี่ยนแปลงในจำนวนผู้ล่าและเหยื่อจะสร้างวิถีโคจรที่บิดเข้าด้านใน ซึ่งสอดคล้องกับความผันผวนที่หน่วงของความอุดมสมบูรณ์ ของเหยื่อและผู้ล่า (รูปที่ 8, A) ในกรณีที่ไอโซไลน์นักล่าตัดกับไอโซไลน์ของเหยื่อในส่วนที่ขึ้น (เช่น ในพื้นที่ ค่าต่ำความหนาแน่นของเหยื่อ) เวกเตอร์สร้างวิถีการคลี่คลายและความผันผวนของจำนวนผู้ล่าและเหยื่อเกิดขึ้นตามลำดับเมื่อแอมพลิจูดเพิ่มขึ้น (รูปที่ 8, ข)หากไอโซไลน์ของนักล่าตัดกับไอโซไลน์ของเหยื่อในบริเวณปลายของมัน เวกเตอร์จะก่อตัวเป็นวงกลมปิด และความผันผวนของจำนวนเหยื่อและผู้ล่านั้นมีลักษณะเฉพาะด้วยแอมพลิจูดและคาบที่มั่นคง (รูปที่ 8, วี)
กล่าวอีกนัยหนึ่ง การแกว่งแบบหน่วงนั้นสอดคล้องกับสถานการณ์ที่ผู้ล่ามีผลกระทบที่เห็นได้ชัดเจนต่อจำนวนเหยื่อที่มีความหนาแน่นสูงมากเท่านั้น (ใกล้กับค่าสูงสุด) และการแกว่งของแอมพลิจูดที่เพิ่มขึ้นเกิดขึ้นเมื่อผู้ล่าสามารถ เพิ่มจำนวนอย่างรวดเร็วแม้จะมีเหยื่อที่มีความหนาแน่นต่ำและมีวิธีทำลายมันอย่างรวดเร็ว ในแบบจำลองเวอร์ชันอื่น Posenzweig และ McArthur แสดงให้เห็นว่าการสั่นของนักล่าและเหยื่อสามารถทำให้เสถียรได้โดยการแนะนำ "ที่หลบภัย" กล่าวคือ บ่งบอกว่าในพื้นที่ที่มีเหยื่อหนาแน่นน้อยจะมีบริเวณที่เหยื่อมีความอุดมสมบูรณ์เพิ่มขึ้นโดยไม่คำนึงถึงจำนวนผู้ล่าที่มีอยู่
ความปรารถนาที่จะสร้างแบบจำลองที่สมจริงยิ่งขึ้นโดยการเพิ่มความซับซ้อนนั้นได้แสดงออกมาในผลงานของไม่เพียงแต่นักทฤษฎีเท่านั้น แต่ยังรวมถึง ผู้ทดลอง โดยเฉพาะอย่างยิ่ง Huffaker ได้รับผลลัพธ์ที่น่าสนใจซึ่งแสดงให้เห็นถึงความเป็นไปได้ของการอยู่ร่วมกันของนักล่าและเหยื่อในโหมดการสั่นโดยใช้ตัวอย่างของไรกินพืชขนาดเล็ก อีโอเตตรานีคัส เซ็กซ์มาคูลาทัสและมีเห็บนักล่ามาโจมตีเขา ไทฟิลโดรมัส ตะวันตก. ส้มที่วางอยู่บนถาดที่มีรู (เช่นเดียวกับที่ใช้เก็บและขนไข่) ถูกนำมาใช้เป็นอาหารของไรที่กินพืชเป็นอาหาร เวอร์ชันดั้งเดิมมี 40 รูบนถาดเดียว บางถาดมีส้ม (ปอกเปลือกบางส่วน) และบางถาดมีลูกบอลยาง เห็บทั้งสองชนิดแพร่พันธุ์ได้เร็วมาก ดังนั้นธรรมชาติของการเปลี่ยนแปลงของประชากรจึงสามารถเปิดเผยได้ในระยะเวลาอันสั้น เมื่อวางไรเพศเมีย 20 ตัวลงบนถาด Huffaker สังเกตเห็นการเติบโตอย่างรวดเร็วของประชากรซึ่งมีความเสถียรที่ระดับ 5-8,000 ตัว (ต่อส้ม) หากนักล่าหลายคนถูกเพิ่มเข้าไปในจำนวนเหยื่อที่เพิ่มขึ้น จำนวนประชากรของเหยื่อกลุ่มหลังก็จะเพิ่มขึ้นอย่างรวดเร็วและตายไปเมื่อเหยื่อทั้งหมดถูกกิน
ด้วยการเพิ่มขนาดของถาดเป็น 120 รู โดยสุ่มส้มแต่ละผลกระจายไปตามลูกบอลยางหลายๆ ลูก Huffaker จึงสามารถยืดเวลาการอยู่ร่วมกันของผู้ล่าและเหยื่อได้ เมื่อปรากฎอัตราส่วนของอัตราการแพร่กระจายของพวกมันมีบทบาทสำคัญในปฏิสัมพันธ์ระหว่างผู้ล่าและเหยื่อ ฮัฟเฟเกอร์แนะนำว่าการทำให้เหยื่อเคลื่อนไหวได้ง่ายขึ้นและการเคลื่อนไหวของนักล่ายากขึ้น จะทำให้ระยะเวลาอยู่ร่วมกันของพวกมันยาวนานขึ้น ในการทำเช่นนี้บนถาด 120 หลุม สุ่มวางส้ม 6 ลูกไว้ท่ามกลางลูกบอลยาง และรอบๆ หลุมที่มีส้ม วาสลีนได้ถูกสร้างขึ้นเพื่อป้องกันไม่ให้ผู้ล่าแพร่กระจาย และหมุดไม้ก็เพื่อความสะดวกในการตั้งถิ่นฐานของเหยื่อ เสริมความแข็งแกร่งบนถาดซึ่งทำหน้าที่เป็น "แผ่นบินขึ้น" สำหรับไรที่กินพืชเป็นอาหาร (ความจริงก็คือสายพันธุ์นี้ผลิตเส้นไหมบาง ๆ และด้วยความช่วยเหลือจากพวกมันสามารถลอยอยู่ในอากาศกระจายไปตามลม) ในแหล่งที่อยู่อาศัยที่ซับซ้อนเช่นนี้ ผู้ล่าและเหยื่ออยู่ร่วมกันเป็นเวลา 8 เดือน แสดงให้เห็นถึงความผันผวนของประชากรครบสามรอบ เงื่อนไขที่สำคัญที่สุดสำหรับการอยู่ร่วมกันนี้มีดังต่อไปนี้: ความหลากหลายของแหล่งที่อยู่อาศัย (ในแง่ของการมีอยู่ของพื้นที่ที่เหมาะสมและไม่เหมาะสมสำหรับเหยื่อที่จะอาศัยอยู่) เช่นเดียวกับความเป็นไปได้ของการย้ายถิ่นของเหยื่อและผู้ล่า (ในขณะที่ยังคงรักษา ข้อได้เปรียบของเหยื่อบางประการในเรื่องความเร็วของกระบวนการนี้) กล่าวอีกนัยหนึ่งผู้ล่าสามารถกำจัดเหยื่อที่สะสมอยู่ในท้องถิ่นได้อย่างสมบูรณ์ แต่เหยื่อบางคนจะมีเวลาในการอพยพและก่อให้เกิดการสะสมในท้องถิ่นอื่น ๆ ไม่ช้าก็เร็วนักล่าก็จะไปถึงการสะสมในท้องถิ่นใหม่ แต่ในระหว่างนี้เหยื่อจะมีเวลาไปตั้งถิ่นฐานในสถานที่อื่น (รวมถึงที่ที่มันเคยอาศัยอยู่มาก่อน แต่ถูกกำจัดออกไปแล้ว)
สิ่งที่คล้ายคลึงกับสิ่งที่ Huffaker สังเกตในการทดลองก็เกิดขึ้นในสภาพธรรมชาติเช่นกัน เช่น ผีเสื้อกลางคืนกระบองเพชร (กระบองเพชร ต้นกระบองเพชร), นำมาออสเตรเลียลดจำนวนกระบองเพชรแพร์เต็มไปด้วยหนามลงอย่างมาก แต่ไม่ได้ทำลายมันอย่างแม่นยำเพราะกระบองเพชรจะแพร่กระจายเร็วขึ้นเล็กน้อย ในสถานที่เหล่านั้นซึ่งกำจัดลูกแพร์เต็มไปด้วยหนามมอดก็จะไม่เกิดขึ้น ดังนั้นเมื่อผ่านไประยะหนึ่งเมื่อลูกแพร์เต็มไปด้วยหนามแทรกซึมมาที่นี่อีกครั้งก็สามารถเติบโตได้ในช่วงระยะเวลาหนึ่งโดยไม่เสี่ยงต่อการถูกมอดทำลาย อย่างไรก็ตามเมื่อเวลาผ่านไปมอดก็ปรากฏขึ้นที่นี่อีกครั้งและเมื่อขยายพันธุ์อย่างรวดเร็วก็ทำลายลูกแพร์เต็มไปด้วยหนาม
เมื่อพูดถึงความผันผวนของนักล่าและเหยื่อ เราไม่สามารถพลาดที่จะพูดถึงการเปลี่ยนแปลงของวัฏจักรของจำนวนกระต่ายและแมวป่าชนิดหนึ่งในแคนาดา ซึ่งติดตามตามสถิติการเก็บเกี่ยวขนสัตว์โดยบริษัท Hudson Bay ตั้งแต่ปลายศตวรรษที่ 18 ถึงต้นศตวรรษที่ 19 ศตวรรษที่ 20. ตัวอย่างนี้มักถูกมองว่าเป็นตัวอย่างคลาสสิกของการแกว่งของเหยื่อระหว่างผู้ล่า แม้ว่าในความเป็นจริงเราจะเห็นเพียงการเติบโตของจำนวนผู้ล่า (แมวป่าชนิดหนึ่ง) ตามการเติบโตของประชากรเหยื่อ (กระต่าย) สำหรับจำนวนกระต่ายที่ลดลงหลังจากการเพิ่มขึ้นแต่ละครั้งนั้นไม่สามารถอธิบายได้ด้วยแรงกดดันที่เพิ่มขึ้นของผู้ล่าเท่านั้น แต่มีความเกี่ยวข้องกับปัจจัยอื่น ๆ ซึ่งเห็นได้ชัดว่าขาดอาหารในฤดูหนาวเป็นหลัก โดยเฉพาะอย่างยิ่ง M. Gilpin ได้ข้อสรุปนี้ ซึ่งพยายามตรวจสอบว่าแบบจำลอง Lotka-Volterra แบบคลาสสิกสามารถอธิบายข้อมูลเหล่านี้ได้หรือไม่ ผลการทดสอบแสดงให้เห็นว่าไม่มีความพอดีที่น่าพอใจกับแบบจำลอง แต่น่าแปลกที่จะดีกว่าหากสลับผู้ล่าและเหยื่อเข้าด้วยกัน เช่น ตีความแมวป่าชนิดหนึ่งว่าเป็น "เหยื่อ" และกระต่ายเป็น "นักล่า" สถานการณ์ที่คล้ายกันนี้สะท้อนให้เห็นในชื่อบทความที่น่าขบขัน (“ กระต่ายกินแมวป่าชนิดหนึ่งหรือไม่?”) ซึ่งโดยพื้นฐานแล้วมีความจริงจังมากและตีพิมพ์ในวารสารวิทยาศาสตร์ที่จริงจัง
หน่วยงานกลางเพื่อการศึกษา
สถาบันการศึกษาของรัฐ
การศึกษาวิชาชีพชั้นสูง
"มหาวิทยาลัยเทคนิคแห่งรัฐ Izhevsk"
คณะคณิตศาสตร์ประยุกต์
ภาควิชา “การสร้างแบบจำลองทางคณิตศาสตร์ของกระบวนการและเทคโนโลยี”
งานหลักสูตร
ในสาขาวิชา “สมการเชิงอนุพันธ์”
เรื่อง: " การวิจัยเชิงคุณภาพโมเดลนักล่า-เหยื่อ"
อีเจฟสค์ 2010
การแนะนำ
1. พารามิเตอร์และสมการพื้นฐานของแบบจำลอง “ผู้ล่า-เหยื่อ”
2.2 โมเดลวอลแตร์ทั่วไปประเภท "นักล่า-เหยื่อ"
3. การประยุกต์ใช้แบบจำลอง “ผู้ล่า-เหยื่อ” ในเชิงปฏิบัติ
บทสรุป
บรรณานุกรม
การแนะนำ
ปัจจุบันปัญหาสิ่งแวดล้อมมีความสำคัญยิ่ง ขั้นตอนสำคัญในการแก้ปัญหาเหล่านี้คือการพัฒนาแบบจำลองทางคณิตศาสตร์ของระบบนิเวศ
ภารกิจหลักประการหนึ่งของระบบนิเวศในปัจจุบันคือการศึกษาโครงสร้างและการทำงานของ ระบบธรรมชาติ, ค้นหารูปแบบทั่วไป คณิตศาสตร์มีอิทธิพลอย่างมากต่อระบบนิเวศ ซึ่งมีส่วนทำให้เกิดระบบนิเวศทางคณิตศาสตร์ โดยเฉพาะอย่างยิ่งในส่วนต่างๆ เช่น ทฤษฎีสมการเชิงอนุพันธ์ ทฤษฎีเสถียรภาพ และทฤษฎีการควบคุมที่เหมาะสมที่สุด
ผลงานชิ้นแรกในสาขานิเวศวิทยาทางคณิตศาสตร์คืองานของ A.D. Lotki (1880 - 1949) ซึ่งเป็นคนแรกที่อธิบายปฏิสัมพันธ์ของประชากรต่างๆ ที่เชื่อมโยงกันด้วยความสัมพันธ์ระหว่างนักล่าและเหยื่อ การมีส่วนร่วมอย่างมากในการศึกษาแบบจำลองนักล่า - เหยื่อทำโดย V. Volterra (1860 - 1940), V.A. Kostitsyn (2426-2506) ปัจจุบันสมการที่อธิบายปฏิสัมพันธ์ของประชากรเรียกว่าสมการ Lotka-Volterra
สมการ Lotka-Volterra อธิบายพลวัตของค่าเฉลี่ย - ขนาดประชากร ในปัจจุบัน บนพื้นฐานของแบบจำลองเหล่านี้ ได้มีการสร้างแบบจำลองทั่วไปของการโต้ตอบของประชากร ซึ่งอธิบายโดยสมการเชิงปริพันธ์-ดิฟเฟอเรนเชียล และกำลังมีการศึกษาแบบจำลองแบบนักล่า-เหยื่อแบบควบคุม
ปัญหาสำคัญประการหนึ่งของนิเวศวิทยาทางคณิตศาสตร์คือปัญหาเสถียรภาพของระบบนิเวศและการจัดการระบบเหล่านี้ การจัดการสามารถดำเนินการได้โดยมีเป้าหมายในการถ่ายโอนระบบจากสถานะเสถียรหนึ่งไปยังอีกสถานะหนึ่ง เพื่อวัตถุประสงค์ในการใช้งานหรือการฟื้นฟู
1. พารามิเตอร์และสมการพื้นฐานของแบบจำลองนักล่า-ไพรเมต
ความพยายามในการสร้างแบบจำลองทางคณิตศาสตร์เกี่ยวกับพลวัตของประชากรทางชีววิทยาและชุมชนแต่ละกลุ่ม รวมถึงประชากรที่มีปฏิสัมพันธ์กัน หลากหลายชนิด, ได้ดำเนินการมาเป็นเวลานาน หนึ่งในแบบจำลองแรกของการเติบโตของประชากรแบบแยกเดี่ยว (2.1) ได้รับการเสนอย้อนกลับไปในปี พ.ศ. 2341 โดย Thomas Malthus:
โมเดลนี้ระบุโดยพารามิเตอร์ต่อไปนี้:
N - ขนาดประชากร
ความแตกต่างระหว่างอัตราการเกิดและอัตราการตาย
เมื่อรวมสมการนี้เข้าด้วยกันเราได้รับ:
![]() , (1.2)
, (1.2)
โดยที่ N(0) คือขนาดประชากรในขณะนี้ t = 0 แน่นอนว่าแบบจำลอง Malthus ที่ > 0 ให้จำนวนที่เพิ่มขึ้นอย่างไม่สิ้นสุด ซึ่งไม่เคยพบเห็นในประชากรตามธรรมชาติ โดยที่ทรัพยากรที่รับประกันการเติบโตนี้จะถูกจำกัดอยู่เสมอ การเปลี่ยนแปลงจำนวนประชากรของพืชและสัตว์ไม่สามารถอธิบายได้ด้วยกฎแมลธัสเซียนง่ายๆ พลวัตการเจริญเติบโตได้รับอิทธิพลจากสาเหตุหลายประการที่สัมพันธ์กัน โดยเฉพาะอย่างยิ่ง การสืบพันธุ์ของแต่ละสายพันธุ์ได้รับการควบคุมและแก้ไขด้วยตนเอง เพื่อให้สายพันธุ์นี้ได้รับการเก็บรักษาไว้ใน กระบวนการวิวัฒนาการ
คำอธิบายทางคณิตศาสตร์ของรูปแบบเหล่านี้ได้รับการจัดการโดยนิเวศวิทยาทางคณิตศาสตร์ - วิทยาศาสตร์เกี่ยวกับความสัมพันธ์ของสิ่งมีชีวิตพืชและสัตว์และชุมชนที่พวกมันก่อตัวขึ้นระหว่างกันและกับสิ่งแวดล้อม
การศึกษาแบบจำลองชุมชนทางชีววิทยาที่จริงจังที่สุด รวมถึงประชากรหลายสายพันธุ์ ดำเนินการโดยนักคณิตศาสตร์ชาวอิตาลี Vito Volterra:
 ,
,
ขนาดประชากรอยู่ที่ไหน
อัตราการเพิ่ม (หรือการตาย) ตามธรรมชาติของประชากร - ค่าสัมประสิทธิ์ของการโต้ตอบระหว่างกัน แบบจำลองนี้อธิบายถึงการต่อสู้ของสายพันธุ์ต่างๆ เพื่อทรัพยากรร่วมกัน หรือปฏิสัมพันธ์ระหว่างนักล่าและเหยื่อ เมื่อสายพันธุ์หนึ่งเป็นอาหารของอีกสายพันธุ์หนึ่ง ทั้งนี้ขึ้นอยู่กับการเลือกค่าสัมประสิทธิ์ หากผลงานของผู้เขียนคนอื่นมุ่งเน้นไปที่การสร้างแบบจำลองต่าง ๆ V. Volterra ได้ทำการศึกษาเชิงลึกเกี่ยวกับแบบจำลองที่สร้างขึ้นของชุมชนทางชีววิทยา ตามที่นักวิทยาศาสตร์หลายคนกล่าวไว้ในหนังสือของ V. Volterra ว่านิเวศวิทยาทางคณิตศาสตร์สมัยใหม่เริ่มต้นขึ้น
2. การวิจัยเชิงคุณภาพแบบจำลอง “ผู้ล่า-เหยื่อ” เบื้องต้น
2.1 แบบจำลองปฏิสัมพันธ์ทางโภชนาการตามประเภท "นักล่า-เหยื่อ"
ให้เราพิจารณาแบบจำลองปฏิสัมพันธ์ทางโภชนาการของประเภท "นักล่า - เหยื่อ" ที่สร้างโดย V. Volterra ให้มีระบบที่ประกอบด้วยสองสายพันธุ์ ชนิดหนึ่งกินอีกชนิดหนึ่ง
ลองพิจารณากรณีที่สัตว์ชนิดหนึ่งเป็นสัตว์นักล่าและอีกสัตว์หนึ่งเป็นเหยื่อ เราจะถือว่าสัตว์นักล่ากินเหยื่อเพียงอย่างเดียวเท่านั้น เรามายอมรับสมมติฐานง่ายๆ ต่อไปนี้:
อัตราการเติบโตของเหยื่อ
อัตราการเติบโตของนักล่า
ขนาดประชากรเหยื่อ
ขนาดประชากรนักล่า
อัตราเหยื่อเพิ่มขึ้นตามธรรมชาติ
อัตราการบริโภคเหยื่อของนักล่า
อัตราการตายของผู้ล่าในกรณีที่ไม่มีเหยื่อ
ค่าสัมประสิทธิ์การ "แปรรูป" ของชีวมวลที่เป็นเหยื่อโดยผู้ล่าให้เป็นชีวมวลของมันเอง
จากนั้นพลวัตของประชากรในระบบนักล่า - เหยื่อจะถูกอธิบายโดยระบบสมการเชิงอนุพันธ์ (2.1):
 (2.1)
(2.1)
โดยที่สัมประสิทธิ์ทั้งหมดเป็นบวกและคงที่
โมเดลก็มี สารละลายสมดุล (2.2):
ตามแบบจำลอง (2.1) สัดส่วนของผู้ล่าใน มวลรวมสัตว์แสดงตามสูตร (2.3):
 (2.3)
(2.3)
การวิเคราะห์ความเสถียรของสถานะสมดุลในส่วนที่เกี่ยวข้องกับการรบกวนเล็กน้อย แสดงให้เห็นว่าจุดเอกพจน์ (2.2) มีความเสถียร "เป็นกลาง" (ของประเภท "ศูนย์กลาง") กล่าวคือ การเบี่ยงเบนใด ๆ จากสมดุลจะไม่ตายไป แต่ถ่ายโอน ระบบเข้าสู่โหมดการสั่นด้วยแอมพลิจูดขึ้นอยู่กับขนาดของสัญญาณรบกวน วิถีของระบบบนระนาบเฟสมีรูปแบบของเส้นโค้งปิดซึ่งอยู่ห่างจากจุดสมดุลต่างๆ (รูปที่ 1)

ข้าว. 1 – ระยะ “ภาพเหมือน” ของระบบ “นักล่า-เหยื่อ” ของโวลแตร์ราแบบคลาสสิก
เมื่อหารสมการแรกของระบบ (2.1) ด้วยสมการที่สอง เราจะได้สมการเชิงอนุพันธ์ (2.4) สำหรับเส้นโค้งบนระนาบเฟส
 (2.4)
(2.4)
เมื่อรวมสมการนี้เข้าด้วยกันเราได้รับ:
![]() (2.5)
(2.5)
โดยที่ค่าคงที่ของการอินทิเกรตคือที่ไหน 
เป็นเรื่องง่ายที่จะแสดงให้เห็นว่าการเคลื่อนที่ของจุดไปตามระนาบเฟสจะเกิดขึ้นในทิศทางเดียวเท่านั้น ในการทำเช่นนี้ จะสะดวกในการแทนที่ฟังก์ชันและโดยการย้ายจุดกำเนิดของพิกัดบนเครื่องบินไปยังจุดที่นิ่ง (2.2) จากนั้นจึงแนะนำพิกัดเชิงขั้ว:
 (2.6)
(2.6)
ในกรณีนี้การแทนที่ค่าของระบบ (2.6) ลงในระบบ (2.1) เราจะได้:
 (2.7)
(2.7)
คูณสมการแรกด้วยและสมการที่สองแล้วบวกเข้าด้วยกัน เราจะได้:
หลังจากการแปลงพีชคณิตที่คล้ายกัน เราจะได้สมการสำหรับ:
ปริมาณดังที่เห็นได้จาก (4.9) จะต้องมากกว่าศูนย์เสมอ จึงไม่เปลี่ยนเครื่องหมาย และการหมุนจะหมุนไปในทิศทางเดียวเสมอ
การรวม (2.9) เราพบช่วงเวลา:
เมื่อมีขนาดเล็ก สมการ (2.8) และ (2.9) จะกลายเป็นสมการวงรี ระยะเวลาการหมุนเวียนในกรณีนี้เท่ากับ:
 (2.11)
(2.11)
ขึ้นอยู่กับช่วงเวลาของการแก้สมการ (2.1) เราสามารถรับผลลัพธ์บางอย่างได้ ในการดำเนินการนี้ ให้เราเป็นตัวแทน (2.1) ในรูปแบบ:
 (2.12)
(2.12)
และบูรณาการในช่วงเวลา:
 (2.13)
(2.13)
เนื่องจากการทดแทนจากและเนื่องจากช่วงเวลาหายไป ค่าเฉลี่ยของช่วงเวลาจึงเท่ากับสถานะคงที่ (2.14):
 (2.14)
(2.14)
สมการที่ง่ายที่สุดของแบบจำลอง "นักล่า-เหยื่อ" (2.1) มีจำนวนดังนี้ ข้อบกพร่องที่สำคัญ. ดังนั้นพวกเขาจึงถือว่าทรัพยากรอาหารไม่ จำกัด สำหรับเหยื่อและการเติบโตของนักล่าอย่างไม่ จำกัด ซึ่งขัดแย้งกับข้อมูลการทดลอง นอกจากนั้น ดังที่เห็นได้จากรูป 1 ไม่มีเส้นโค้งเฟสใดที่แตกต่างจากมุมมองของความเสถียร เมื่อมีอิทธิพลรบกวนแม้แต่น้อย วิถีของระบบจะเคลื่อนตัวออกจากตำแหน่งสมดุลมากขึ้นเรื่อยๆ แอมพลิจูดของการแกว่งจะเพิ่มขึ้น และระบบจะพังทลายลงอย่างรวดเร็ว
แม้จะมีข้อบกพร่องของแบบจำลอง (2.1) แนวคิดเกี่ยวกับลักษณะการแกว่งพื้นฐานของพลวัตของระบบ” นักล่าเหยื่อ» แพร่หลายในระบบนิเวศ ปฏิสัมพันธ์ระหว่างผู้ล่าและเหยื่อใช้เพื่ออธิบายปรากฏการณ์ต่างๆ เช่น ความผันผวนของจำนวนสัตว์นักล่าและสัตว์สงบในพื้นที่ประมง ความผันผวนของจำนวนปลา แมลง ฯลฯ อันที่จริง ความผันผวนของจำนวนอาจเกิดจากสาเหตุอื่น
ให้เราสมมติว่าในระบบนักล่า - เหยื่อมีการทำลายล้างของบุคคลทั้งสองสายพันธุ์และพิจารณาคำถามว่าการทำลายของแต่ละบุคคลส่งผลกระทบต่อค่าเฉลี่ยของตัวเลขของพวกเขาอย่างไรหากดำเนินการตามสัดส่วนของจำนวนนี้ด้วยสัดส่วน ค่าสัมประสิทธิ์และตามลำดับสำหรับเหยื่อและผู้ล่า เมื่อคำนึงถึงสมมติฐานแล้ว ให้เขียนระบบสมการ (2.1) ใหม่ในรูปแบบ:
 (2.15)
(2.15)
สมมติว่า นั่นคือสัมประสิทธิ์การกำจัดเหยื่อน้อยกว่าค่าสัมประสิทธิ์การเพิ่มขึ้นตามธรรมชาติ ในกรณีนี้ จะสังเกตความผันผวนของตัวเลขเป็นระยะด้วย มาคำนวณตัวเลขเฉลี่ยกัน:
 (2.16)
(2.16)
ดังนั้น ถ้า ขนาดประชากรโดยเฉลี่ยของเหยื่อเพิ่มขึ้น และผู้ล่าก็ลดลง
ให้เราพิจารณากรณีที่ค่าสัมประสิทธิ์การกำจัดเหยื่อมากกว่าค่าสัมประสิทธิ์การเพิ่มขึ้นตามธรรมชาตินั่นคือ ในกรณีนี้ ![]() สำหรับใดๆ และ ดังนั้น การแก้สมการแรก (2.15) จึงถูกจำกัดขอบเขตจากด้านบนด้วยฟังก์ชันลดลงแบบเอกซ์โปเนนเชียล
สำหรับใดๆ และ ดังนั้น การแก้สมการแรก (2.15) จึงถูกจำกัดขอบเขตจากด้านบนด้วยฟังก์ชันลดลงแบบเอกซ์โปเนนเชียล ![]() , ฉันกิน .
, ฉันกิน .
เริ่มต้นจากช่วงเวลาหนึ่ง t ซึ่ง ณ เวลานั้น การแก้สมการที่สอง (2.15) ก็เริ่มลดลงและมีแนวโน้มเป็นศูนย์เช่นกัน ดังนั้นในกรณีที่ทั้งสองสายพันธุ์หายไป
2.1 โมเดลวอลแตร์ทั่วไปประเภท "นักล่า-เหยื่อ"
โดยธรรมชาติแล้วโมเดลแรกของ V. Volterra ไม่สามารถสะท้อนทุกแง่มุมของการมีปฏิสัมพันธ์ในระบบล่าเหยื่อได้เนื่องจากพวกมันถูกทำให้ง่ายขึ้นมากเมื่อเทียบกับสภาพจริง ตัวอย่างเช่น หากจำนวนผู้ล่าเป็นศูนย์ ดังนั้นจากสมการ (1.4) จะตามมาว่าจำนวนเหยื่อเพิ่มขึ้นอย่างไม่มีกำหนด ซึ่งไม่เป็นความจริง อย่างไรก็ตาม คุณค่าของแบบจำลองเหล่านี้อยู่ที่ความจริงที่ว่าแบบจำลองเหล่านี้เป็นพื้นฐานที่ทำให้นิเวศวิทยาทางคณิตศาสตร์เริ่มพัฒนาอย่างรวดเร็ว
มีการศึกษาจำนวนมากเกี่ยวกับการดัดแปลงระบบนักล่าและเหยื่อที่หลากหลาย โดยมีการสร้างแบบจำลองทั่วไปที่คำนึงถึงสถานการณ์จริงในธรรมชาติในระดับหนึ่งหรืออย่างอื่น
ในปี พ.ศ. 2479 A.N. Kolmogorov เสนอให้ใช้เหยื่อนักล่าเพื่ออธิบายพลวัตของระบบ ระบบต่อไปนี้สมการ:
 , (2.17)
, (2.17)
โดยที่มันจะลดลงตามจำนวนผู้ล่าที่เพิ่มขึ้น และเพิ่มขึ้นตามจำนวนเหยื่อที่เพิ่มขึ้น
ระบบสมการเชิงอนุพันธ์นี้เนื่องจากความทั่วไปที่เพียงพอทำให้สามารถคำนึงถึงพฤติกรรมที่แท้จริงของประชากรและในเวลาเดียวกันก็ดำเนินการ การวิเคราะห์เชิงคุณภาพการตัดสินใจของเธอ
ต่อมาในงานของเขา Kolmogorov สำรวจอย่างละเอียดน้อยลง รุ่นทั่วไป:
 (2.18)
(2.18)
กรณีพิเศษต่างๆ ของระบบสมการเชิงอนุพันธ์ (2.18) ได้รับการศึกษาโดยผู้เขียนหลายคน ตารางแสดงกรณีพิเศษต่างๆ ของฟังก์ชัน , ,
ตารางที่ 1 - แบบจำลองชุมชนนักล่า-เหยื่อต่างๆ
| ผู้เขียน | |||
| โวลแตร์รา ลอตก้า | |||
| เกาส์ | |||
| พีสโลว์ | |||
| โฮลิ่ง | |||
| อิฟเลฟ | |||
| โรยามะ | |||
| ชิมาซึ | |||
| อาจ |
เหยื่อนักล่าแบบจำลองทางคณิตศาสตร์
3. การนำไปปฏิบัติจริงของแบบจำลองผู้ล่า-เหยื่อ
ลองพิจารณาแบบจำลองทางคณิตศาสตร์ของการอยู่ร่วมกันของสองสายพันธุ์ทางชีววิทยา (ประชากร) ประเภท "นักล่า - เหยื่อ" ที่เรียกว่าแบบจำลอง Volterra - Lotka
ให้สอง สายพันธุ์ทางชีวภาพอยู่ด้วยกันในสภาพแวดล้อมที่โดดเดี่ยว สิ่งแวดล้อมอยู่นิ่งและให้ทุกสิ่งที่จำเป็นสำหรับชีวิตในปริมาณไม่จำกัดแก่หนึ่งในสายพันธุ์ ซึ่งเราจะเรียกว่าเหยื่อ อีกสายพันธุ์หนึ่งซึ่งเป็นนักล่าก็อยู่ในสภาพนิ่งเช่นกัน แต่จะกินเฉพาะบุคคลในสายพันธุ์แรกเท่านั้น สิ่งเหล่านี้อาจเป็นปลาคาร์พและหอกไม้กางเขน กระต่ายและหมาป่า หนูและสุนัขจิ้งจอก จุลินทรีย์และแอนติบอดี ฯลฯ เพื่อความชัดเจน เราจะเรียกพวกมันว่าปลาคาร์พและหอกไม้กางเขน
ระบุพารามิเตอร์เริ่มต้นต่อไปนี้:
เมื่อเวลาผ่านไป จำนวนปลาคาร์พ crucian และหอกเปลี่ยนแปลงไป แต่เนื่องจากมีปลาจำนวนมากในบ่อ เราจะไม่แยกความแตกต่างระหว่างปลาคาร์พ crucian 1,020 ตัวและ 1,021 ดังนั้นเราจะถือว่าพวกมันเป็นฟังก์ชันต่อเนื่องของเวลา t ด้วย เราจะเรียกคู่ของตัวเลข (,) ว่าสถานะของโมเดล
เห็นได้ชัดว่าธรรมชาติของการเปลี่ยนแปลงในสถานะ (,) ถูกกำหนดโดยค่าของพารามิเตอร์ ด้วยการเปลี่ยนพารามิเตอร์และการแก้ระบบสมการของแบบจำลอง ทำให้สามารถศึกษารูปแบบของการเปลี่ยนแปลงสถานะของระบบนิเวศเมื่อเวลาผ่านไปได้
ในระบบนิเวศ อัตราการเปลี่ยนแปลงจำนวนของแต่ละสายพันธุ์จะถือเป็นสัดส่วนกับจำนวนของมันด้วย แต่จะมีค่าสัมประสิทธิ์ที่ขึ้นอยู่กับจำนวนบุคคลในสายพันธุ์อื่นเท่านั้น ดังนั้นสำหรับปลาคาร์พ crucian ค่าสัมประสิทธิ์นี้จะลดลงตามจำนวนหอกที่เพิ่มขึ้น และสำหรับหอกนั้นจะเพิ่มขึ้นตามจำนวนปลาคาร์พ crucian ที่เพิ่มขึ้น เราจะถือว่าการพึ่งพานี้เป็นเส้นตรงด้วย จากนั้นเราจะได้ระบบสมการเชิงอนุพันธ์สองแบบ:

ระบบสมการนี้เรียกว่าแบบจำลองโวลแตร์รา-ลอตกา ค่าสัมประสิทธิ์ตัวเลข , เรียกว่าพารามิเตอร์แบบจำลอง เห็นได้ชัดว่าธรรมชาติของการเปลี่ยนแปลงในสถานะ (,) ถูกกำหนดโดยค่าของพารามิเตอร์ ด้วยการเปลี่ยนพารามิเตอร์เหล่านี้และการแก้ระบบสมการแบบจำลอง ทำให้สามารถศึกษารูปแบบของการเปลี่ยนแปลงในสถานะของระบบนิเวศได้
ลองรวมสมการทั้งสองของระบบเทียบกับ t ซึ่งจะเปลี่ยนจากช่วงเวลาเริ่มต้นเป็น โดยที่ T คือช่วงเวลาที่การเปลี่ยนแปลงเกิดขึ้นในระบบนิเวศ ในกรณีของเราให้ระยะเวลาเป็น 1 ปี จากนั้นระบบจะใช้แบบฟอร์มต่อไปนี้:
 ;
;
 ;
;
เมื่อหา = และ = และนำพจน์ที่คล้ายกันมา เราจะได้ระบบที่ประกอบด้วยสองสมการ:

โดยการแทนที่ข้อมูลเริ่มต้นในระบบผลลัพธ์ เราจะได้จำนวนประชากรของปลาหอกและปลาคาร์พ crucian ในทะเลสาบหลังจากผ่านไปหนึ่งปี:


